
腦電波
維基百科嘅醫學類文章只係提供參考,並唔係醫療意見。 閣下如果有任何關於健康嘅問題,請諮詢專業醫護人員。 |


舉附圖做簡單例子:想像有一柞神經細胞喺度處理資訊;附圖嘅 Y 軸代表微電壓,而 X 軸代表時間 ,上圖喺每個時間點,都有若干吓神經細胞射訊號引致嘅微電壓(每粒藍色點代表一吓);而附圖下圖代表咗成柞神經細胞嘅整體活動-即係攞每一個時間點,將嗰個時間點嘅微電壓加埋一齊;由下圖嗰度睇得出,柞神經細胞嘅整體活動有週期性。

腦電波研究歷史唔耐,但就好多姿多采:最早嘅腦電波研究係喺 1924 年由德國神經科學家漢斯伯格(Hans Berger)做嘅[4];喺打後嗰半個世紀內,神經科學界都有對脊椎動物嘅神經細胞嘅呢啲振盪作出研究,但就唔多明呢啲振盪係做乜嘢功能嘅[5][6];不過自從喺廿一世紀起,科學界有咗先進嘅神經成像技術,可以攞到高解像度嘅腦部影像,加深咗人類對腦電波嘅理解,而且呢啲研究仲俾人應用落去腦機埠(BCI)等嘅技術嗰度[7][8]。
基礎
[編輯]

神經細胞
[編輯]神經細胞(neuron)係一種與別不同嘅細胞,佈滿動物嘅神經系統:神經細胞分好多種,每種都會對某啲特定類型嘅刺激有反應;當一粒神經細胞受到佢相應嗰種刺激嗰陣,會沿由佢細胞體伸出嚟嗰條軸突(axon)射出電脈衝(動作電位);而好多神經細胞喺受到第啲神經細胞射嘅電脈衝刺激嗰時,又會射新嘅電脈衝-所以當一粒神經細胞受刺激而射電脈衝嗰陣時,好多時就會引致一連串嘅電脈衝,令到訊號沿住神經系統傳開去,一路上到腦[9][10];舉個例說明,喺啲感覺器官嗰度嘅感覺神經細胞(sensory neuron)喺受到光、聲、氣味同溫度變化等嘅嘢刺激到嘅時候,都會射電脈衝,而如果負責感應光嗰啲感覺神經細胞唔射電脈衝,個腦就知道外界冇光[11]。
神經細胞嘅呢啲活動會造成腦電波:一隻動物嘅神經系統喺任何一個時間點-就算佢瞓緊覺都好-都會有大量電活動,涉及電訊號喺神經系統當中傳嚟傳去,而電嘅流動會引致電壓改變(可以睇古典電磁學);只要有方法量度,研究者就可以觀察到腦好多區域嘅微電壓[註 1]會各自週期性(periodic;指每隔一段恆定時間就重複一次)噉上上落落[註 2]-呢啲週期性嘅上上落落就係所謂嘅腦電波[9]。
量度方法
[編輯]腦電波可以用腦電圖(electroencephalogram;簡稱「EEG」)嘅方法量度。簡單講,腦電圖做嘅係喺個頭殼表面或者裏面擺一啲電極[註 3][12],用電極量度腦唔同區域喺唔同時間點嘅微電壓;呢啲數據可以用時頻分析(time-frequency analysis)嘅方法處理(睇埋傅立葉變換)[13][14],得出每個量度咗嘅區域喺某段時間內嘅腦電波嘅頻率、波幅同埋相位等嘅特性[15][16]。
觀察腦電波嘅呢啲特性對於神經科學同心理學上嘅研究好有用:神經系統入面嘅電活動源於神經細胞之間傳嘅資訊,所以腦電波嘅特性能夠反映啲細胞傳緊乜嘢資訊;例如大規模(指橫跨多個腦區)嘅腦電波波幅改變可以表示嗰啲腦區嘅同步程度有所改變,可能表示佢哋活躍咗或者變得冇咁活躍;跟手神經科學家同心理學家就可以靠睇吓呢啲改變同邊啲外界刺激有統計相關(correlation),並且得知個腦邊啲區域係負責乜嘢功能嘅,例如如果腦區 A 喺一個人做一啲要求處理視覺資訊嘅工作嗰陣零舍活躍,噉腦區 A 就應該係做一啲同視覺有關嘅功能[17][18]。
數學性啲噉講,要研究腦電波,神經科學家多數會將個腦當成一個動態系統(dynamic system)噉嚟睇。噉講即係話,研究者會考慮個腦入面嗰啲物理量(微電壓等)隨時間嘅變化,用一啲微分方程嚟模擬研究緊嘅神經系統嘅唔同細部份各自嘅活動,以及係呢啲各自嘅活動點樣加埋一齊俾出所攞到嘅數據顯示嘅活動。呢啲方程如果成功模擬到所得到嘅數據嘅話,啲研究者就能夠當呢啲方程做「能夠準確描述啲腦電波嘅數學模型」,並且睇吓方程當中嘅變數分別同邊啲外界刺激有統計相關[19]。
分析層次
[編輯]
腦電波基本上可以由好幾個層次嚟分析:微層次(micro-scale;單一一粒神經細胞)、中層次(meso-scale;一細柞神經細胞)同埋宏層次(macro-scale;比較唔同腦區)[16]。
微層次
[編輯]動作電位(action potential)係腦電波嘅最基本部份:動作電位係神經細胞射嘅電脈衝訊號,當一個動作電位沿住粒神經細胞嘅軸突傳去下粒神經細胞嗰度嗰陣時,條軸突表面浸細胞膜會有極性上嘅改變。當浸膜某個位嘅電位超過咗某個門檻值嗰陣,佢表面嗰啲鈉(Na+)同鉀(K+)離子門會打開,等呢啲(帶電荷嘅)離子過到,例如係鈉離子門噉,佢會喺動作電位開始嗰時打開,令到鈉離子入去條軸突入面,引起去極化效應,跟手鉀離子門會打開,令到鉀離子離開條軸突,令到條軸突重新極化。呢種作用令到下一個位置再受刺激並且做呢個過程,於是動作電位就會沿住條軸突傳落去。最後去到軸突嘅尾嗰度,下一粒神經細胞會收到呢個訊號,並且將個資訊再傳落去[9][20]。
神經細胞仲曉產生連串動作電位(spike train),呢啲連串嘅動作電位係神經編碼嘅主要基礎-即係話呢啲連串嘅動作電位當中帶有神經細胞所傳嘅資訊。舉個例說明:當腦嘅某一粒神經細胞由對眼嗰度嗰啲感光細胞嗰度收到動作電位訊號(呢啲訊號可能包括咗「睇到邊點有啲乜嘢顏色」等嘅資訊),佢浸細胞膜嗰度就會有電壓改變,而如果呢個電壓改變大過某個門檻值,噉個電壓改變就會會令粒神經細胞射一個新嘅動作電位(個門檻值係幾多視乎好多因素而定),等下一粒神經細胞能夠知道佢所收到嘅資訊,如是者,資訊就能夠喺成個神經網絡嗰度傳開去。單一一粒神經細胞嘅呢啲振盪活動可以透過睇佢膜電位嘅上上落落嚟觀察得到[18]。一連串嘅動作電位可以以好多唔同嘅規律出現,有啲係有節奏噉上上落落嘅,又有啲會爆發性噉上落跟手話咁快消失嘅[21]。
腦電波好大程度上係眾多神經細胞所產生嘅動作電位嘅總和。一粒神經細胞所產生嘅動作電位極之微細,難以真接量度-據估計,一粒神經細胞嘅動作電位頂櫳都係得嗰幾十毫伏特(millivolt;1 毫伏特相當於 1000 份之 1 伏特)左右咁大[22],但就算係一隻昆蟲嘅腦都可以有成幾十萬粒神經細胞,所以呢啲活動嘅總和可以明顯噉用架生探測得到[23]。
中層次
[編輯]一柞神經細胞都可以產生振盪性(上上落落)嘅活動。神經細胞之間會有突觸(synapse)連住,呢啲結構令到神經細胞能夠互傳資訊,而透過突觸嘅互動,神經細胞可以同步(synchronized)噉活動-當一柞神經細胞唔同步活動嗰陣,佢哋喺每個時間點嘅總活動(即係啲動作電位加埋一齊)會唔定,但當佢哋同步一致噉射動作電位嗰時,佢哋嘅總活動就會變得有節奏,喺同步嘅情況下,如果粒粒神經細胞嘅活動都係週期性噉上上落落嘅話,佢哋會喺差唔多同一時間達到最高點,又會喺差唔多同一時間達到最低點,令到佢哋個總活動成為一個大波幅嘅振盪性活動。一大柞同步神經細胞嘅總體活動就會產生夠大嘅電壓改變,可以喺頭皮嗰度用電極量度得到(但呢啲電壓改變都得嗰幾十毫伏特咁大)。腦電圖或者腦磁圖呢啲神經科學上量度腦電波嘅方法多數都係反映緊數以百萬計嘅神經細胞嘅同步活動嘅[24][25]。


以上面嘅圖解說明:Y 軸代表電壓,X 軸代表時間,左圖同右圖下面兩條幼線代表個別神經細胞嘅活動,而最上面嘅粗線代表兩條幼線嘅活動加埋-由左圖睇得出,當兩粒細胞嘅活動完全唔同步(即係一粒細胞去到最高點嗰陣,另外嗰粒啱啱處於最低點),佢哋嘅活動總和會係 0;而右圖就顯示,如果兩粒細胞嘅活動完全同步(兩粒同時去到最高點,又同時去到最低點),佢哋嘅活動總和就會到達最大數值。
是但搵柞神經細胞,佢裏面嗰啲神經細胞好少可會真係做到完全同步,多數都只係可以做到差唔多完全同步(相位差異極細,但唔等如 0)。當一柞神經細胞要同步活動嗰陣時,佢哋喺每個時間點射動作電位嘅機會率會受調控,令到佢哋變得更大機會會一齊射動作電位,產生大嘅總體活動[21]。神經細胞網絡可以內源-即係唔靠外界刺激-噉用自身內部刺激性神經細胞同壓抑性神經細胞之間嘅互動嚟同步化,當中壓抑性(inhibitory)神經細胞零舍重要-呢啲神經細胞曉壓抑第啲神經細胞嘅活動,所以做到唔俾周圍嘅神經細胞喺唔同步嘅時間點射動作電位[26]。
宏層次
[編輯]腦電波亦都可以嚟自唔同腦區之間嘅互動。是但搵兩個腦區,兩者之間都可以有具有相向嘅相配性(bidirectionally coupled),令兩者之間嘅連繫形成到循環嘅回輸-噉講意思即係話,兩個腦區之間能夠互相影響,例如係丘腦(thalamus;喺個腦深處負責將啲感官資訊傳去大腦皮層各區嘅結構)同大腦皮層(cerebral cortex;人腦最外面嗰浸神經組織)之間嘅連繫噉[27],喺正常運作嗰陣,丘腦嘅腦電波會同大腦皮層某啲區嘅腦電波同步化,而呢個過程會令資訊喺兩個腦區之間互傳。順帶一提,呢種活動喺 alpha 波嘅產生當中扮演咗好緊要嘅角色[28][29]。
腦電頻寬
[編輯]喺攞到腦電圖數據之後,就要郁手做分析。腦電圖會俾嘅原始數據(raw data)包含兩樣重要嘅資訊:
- 一大柞嘅 ; 係指(頭殼上嘅)位置,而 係指時間, 係頭殼位置 喺時間點 嘅微電壓值[30];睇埋時間序列(time series)。
- 一般嚟講,要做腦電圖分析仲需要能夠描述實驗刺激(experimental stimulus)出現同消失嘅時間點:舉個例說明,假設家陣研究者想知做唔同嘅動作會點樣影響運動皮層(motor cortex;大腦皮層主管身體郁動嗰忽)嘅電活動,於是佢哋就一路量度受試者嘅腦電活動,一路要求佢哋做唔同動作,然後再比較喺唔同動作之下,受試者嘅腦電活動有乜嘢分別;所以除咗一大柞 之外,佢哋嘅數據庫當中一定要有埋一大柞「喺時間點 同 之間,受試者做緊邊個動作」(實驗刺激嘅時間點)先至有得做解答佢哋研究問題必要嘅分析[31]。





其中一種常見嘅腦電圖分析方法係睇吓幅腦電圖入面嗰啲腦電波:喺一個正常行緊嘅人腦入面,有數以百億計嘅神經細胞[註 4]無時無刻喺度以唔同嘅頻率(frequency)射訊號,當中有啲神經細胞會以 5 Hz 嘅頻率射,成一個 5 Hz 嘅波動,有啲會以 15 Hz 嘅頻率射,成一個 15 Hz 嘅波動... 如此類推(可以睇埋波動)。呢啲活動嘅總和形成腦電圖入面條線嗰啲上上落落,數學性噉講,即係話時間點 嘅微電壓 可以用類似噉嘅式表示[32][33]:

- ;

當中 係一個波嘅最大振幅, 反映頻率,而 就係相位; 反映嗰個波喺時間點 嘅振幅,而 就表示唔同頻率嘅波喺 嘅振幅冚唪唥加埋。喺攞到條腦電圖線之後,研究者可以用傅立葉變換(Fourier transform)等嘅數學方法,將每個上落嘅腦電數據拆開,變成每個組成條線嘅波嘅波長-估計每個頻率嘅腦電波嘅波幅同相位[32][34]。




腦電波可以按照佢哋嘅頻率嚟分類,頻率係指個波會喺一秒之內幾多次由一個最高點去到第個最高點,神經科學界最早發現嘅腦電波係 alpha 波(頻率 7.5 至 12.5 Hz)[35],呢種腦電波可以喺個人醒緊但係放鬆嘅狀態之下響佢嘅枕葉(occipital lobe;大腦皮層最近後尾枕嗰忽,專官視覺資訊)嗰度探測得到,而且喺個人合埋眼嗰陣會變強[36]。除咗 alpha 之外,仲有 delta(1 至 4 Hz)、theta(4 至 8 Hz)、beta(13 至 30 Hz)、同 gamma(30 Hz 以上)呀噉。普遍嚟講,一個腦區嘅腦電波頻率愈高就表示個腦區嗰啲神經細胞愈係射訊號射得好密,愈係表示個腦區好忙噉做緊嘢,例如係一個正常嘅大人喺瞓覺瞓得最深嗰啲階段個腦會由比較低頻嘅 delta 波同 theta 波主導住[37],而喺醒住做緊認知作業嗰陣,個腦就好多時會出現好似 gamma 波呢啲高頻嘅腦電波[38][39]。
腦電波分類
[編輯]| 頻寬 | 頻率(Hz) | 位置 | 正常嗰陣出現喺 | 同邊啲病有關 |
|---|---|---|---|---|
| Delta | < 4 | 大人係近額頭,而細路係近後尾枕。 | ||
| Theta | 4-7 | 喺同個人做緊嗰樣嘢冇關嘅腦區域會搵到[46]。 |
| |
| Alpha | 8-15 | 腦比較近後尾枕嘅區域,喺比較主導性嗰一邊腦會勁啲-即係 alpha 波喺左㕭嘅人入面會喺右腦勁啲,而反之亦然;喺休止嗰陣 alpha 波喺個腦中央地帶(C3-C4)最勁。 | ||
| Beta | 16-31 | 腦嘅左右兩邊都有,大致上平衡分佈,尤其係喺近額頭嘅區域。 | ||
| Gamma | > 32 | 體覺皮層(somatosensory cortex) | ||
| Mu | 8-12 | 感覺運動皮層(sensorimotor cortex) |
大體上嚟講,一個人嘅腦整體上由以上邊一個頻寬嘅電活動主導同以下因素有關[61]:
- 年紀:年紀愈細嘅人,佢個腦嘅正常活動就愈偏向低頻,例如出咗世冇幾耐嘅臊孲仔嘅腦喺多數時間都係由最低頻最慢嘅 delta 波嚟主導嘅,但呢種由 delta 波主導嘅狀態喺正常健康嘅大人身上就通常要係喺個人瞓得好淰嗰陣先至會出現[42]。
- 精神狀態:一個人愈係唔清醒,佢個腦嘅正常活動就愈偏向低頻;當一個大人或者青少年覺得攰開始瞌眼瞓嗰陣,佢個腦嘅 theta 波-第二低頻嘅波-強度會提升,而 alpha 波強度會下降,而當一個人瞓緊覺嗰陣,個腦幾乎唔會點有高頻啲(beta 同 gamma 等)嘅腦電波,而且瞓得愈淰個,腦就偏向俾愈低頻嘅波主導。
- 麻醉:一個人受咗麻醉之後嘅腦電狀態同用咗邊種麻醉藥有關;例如用氟烷(halothane)嗰類藥麻醉嘅病人個腦會俾 alpha 同低頻 beta 嘅腦電波主導[62]。
順帶一提,以上呢套腦電活動分類法係舊陣時(廿世紀中)嘅做法,因為嗰陣時啲腦電圖係用紙記低嘅,啲研究員要靠人手數張紙上面有幾多個波形-淨係研究到頻率係整數嘅腦電活動。呢一點搞到個分類法有啲窿窿罅罅位冇咗分類-好似係 theta 同 alpha 之間嘅 7.000... 到 7.999... Hz 噉。於是有人整咗個改良版[63]:
| 頻寬 | 頻率(Hz) |
|---|---|
| Delta | < 4 |
| Theta | ≥ 4 and < 8 |
| Alpha | ≥ 8 and < 14 |
| Beta | ≥ 14 |

-
 Theta 波(Theta wave,取自希臘字母 Θ )
Theta 波(Theta wave,取自希臘字母 Θ )

-
 Alpha 波(Alpha wave,取自希臘字母 Α )
Alpha 波(Alpha wave,取自希臘字母 Α )

-
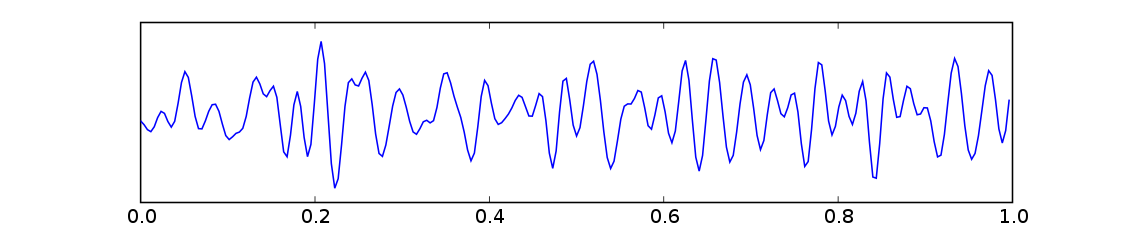 Beta 波(Beta wave,取自希臘字母 Β)
Beta 波(Beta wave,取自希臘字母 Β)

-
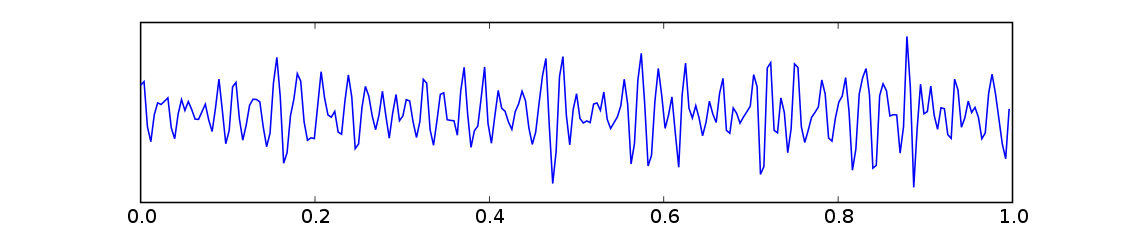 Gamma 波(Gamma wave,取自希臘字母 Γ)
Gamma 波(Gamma wave,取自希臘字母 Γ)

反應方式
[編輯]神經系統嘅神經振盪對外界刺激嘅反應可以大致上分做幾種:無論係單一一粒神經細胞定係一群神經細胞,都曉自發噉產生振盪性嘅活動,而且佢哋仲會對感知輸入(指感知資訊)或者運動輸出(指個腦傳去指揮身體嘅資訊)有振盪性嘅反應;某啲類型嘅神經細胞會無論有冇輸入都照射動作電位,所以一個受試者冇特別做啲乜嘢工作嗰陣嘅腦活動反映咗所謂嘅休息狀態活動(resting-state activity);呢啲活動通常都會喺個人受到外界刺激或者想做某啲郁動嗰陣時有所改變,而改變嘅規律主要可以分做幾大類[16][64]。
| 振盪性嘅反應 | ||||||||
|---|---|---|---|---|---|---|---|---|
|






- 自發活動(spontaneous activity):指個腦喺冇俾外界要求佢做啲乜嘢作業嗰陣時嘅活動,所以又叫「休息狀態活動」[65][66],相對於受觸發嘅活動(induced activity),即係指受外界刺激或者運動輸出而起嘅腦活動[67]。一般嚟講,如果唔係做緊一啲有關自發活動嘅研究,做腦電研究嘅科學家都會將自發活動當做雜音噉嚟睇;雖然係噉,廿一世紀初嘅神經科學界已知,自發活動有相當嘅重要性,喺感知等嘅認知功能上扮演咗一定嘅角色-有腦電圖研究顯示,視覺感知係由皮層神經振盪嘅相位同波幅共同話事嘅,例如喺視覺刺激到達嗰一刻嘅自發 alpha 波活動嘅相位同波幅能夠預測一個受試者會唔會感知得到嗰個刺激[68],即係話自發活動會影響一個人嘅警覺(alertness),而呢樣嘢會影響佢能唔能夠成功感知刺激[69][70]。
- 頻率反應(frequency response):指一粒或者一群神經細胞以改變神經振盪頻率嘅形式嚟對外界輸入俾反應;例如喺感官系統入面,通常當一個外界刺激嘅強度愈高-意思即係個刺激每單位面積嘅功率有幾高,強度高嘅光會掁眼,而強度高嘅聲會響-俾佢刺激到嗰啲神經細胞嘅發射頻率就會愈高,雖然呢個關係通常都唔係線性(linear,指單純嘅正比)嘅[71];頻率改變嘅現象又可以係由個腦嘅內部產生,並且同所跟住產生嘅郁動有關,例如某啲腦區嘅神經振盪頻率俾科學家發現咗會同嗰隻動物嘅行路速度有相關[72]。
- 波幅反應(amplitude response):個腦喺隻動物受到某啲外界刺激嗰陣,有可能會改變持續性嘅神經振盪嘅波幅;例如研究發現,當個腦要做一啲有返咁上下吃力嘅認知作業(好似係喺心入面想像一件物件)嗰陣時,Gamma 波嘅波幅會上升;呢啲反應喺相位上可能唔同步,所以喺神經科學家睇一幅腦電圖嗰時(腦電圖反映嘅係整體活動)會因為呢啲反應互相抵消而睇唔到佢哋,所以要分析呢啲數據實要用到時頻分析先至得[73]。
- 相位重置(phase resetting):指一粒或者一群神經細胞重設佢哋嘅振盪嘅相位(phase)[74];相位重置呢家嘢喺單一神經細胞當中成日都會發生,啲動作電位嘅定時會受到粒神經細胞所受嘅輸入所影響,例如有啲神經細胞一定要喺輸入到達後嘅幾多幾多毫秒先會射動作電位[18],喺神經細胞群嗰度都會見到。相位重置對於神經細胞之間嘅同步嚟講係不可或缺嘅[39][75]。
- 相位重置亦都幫到手研究誘發活動(evoked activity):喺腦電圖等嘅研究領域嗰度,誘發活動係指一啲直接由外界刺激所引起嘅腦活動,例子可以睇事件相關電位(event-related potential);喺科研上,神經科學家可以喺一場實驗當中重複噉做好多次試驗(trial),喺每場試驗入面都要個受試者接觸個外界刺激一次,並且一路監察住個受試者嘅神經振盪活動,最後喺個試驗重複咗(例如)50 次之後就將嗰 50 次試驗嘅腦電波數據計平均數,得出一幅冇嗮隨機變化嘅數據圖。最後得出嘅數據就會係反映咗由個外界刺激引起嘅活動。誘發活動本質上就係指喺外界刺激到達咗若干毫秒之後嘅活動,所以相位重置理論上會係誘發活動嘅主要產生方法[76][77]。
數學模型
[編輯]運算神經科學(computational neuroscience)係指用數學模型嚟模擬神經系統嘅運作:科學一般都追求精確噉描述研究緊嘅現象,而數學符號唔似得自然語言(指廣東話同英文等日常講嘢用嘅語言)噉有咁多歧義嘅問題;原則上,神經振盪同一般嘅物理現象一樣,可以用數學模型嚟描述,而物理學上不嬲都有多種方程式係可以攞嚟模擬振盪性嘅活動嘅,例如係諧振子(harmonic oscillator)噉:諧振子係指(如果冇任何外力)以簡諧運動(simple harmonic motion)振盪嘅物體,喺大自然當中好常見-包括咗聲波、鐘擺嘅運動同埋多種形式嘅震動,而研究表明咗,由雜音帶動嘅諧振子模型能夠相當準確噉模擬到醒緊嘅 alpha 波同埋瞓覺嗰陣嘅慢波;又有啲腦電圖數據比較適合用諧振子嘅變種模型嚟模擬[78][79]。
單一細胞模型
[編輯]
神經科學家可以用數學模型嚟描述神經細胞嘅各種特質,並以一定嘅準確度描述同預測粒神經細胞嘅生物過程。基於電導模型(conductance-based model)係最成功同廣為人採用嘅神經細胞模型,個模型建基於由魷魚巨型軸突(Squid giant axon)嗰度得到嘅數據,用一柞非線性嘅微分方程嚟模擬神經細胞嘅電特性,尤其係動作電位嘅產生同傳播。基於電導模型所涉及嘅數學相當深,所以有科學家嘗試將佢簡化,等其他科學家有得同時模擬大量神經細胞嘅活動。例如軒馬殊-蘿絲模型(Hindmarsh-Rose model)就係基於電導模型嘅其中一個簡化版,軒馬殊-蘿絲模型用咗以下幾條微分方程嚟模擬神經細胞嘅活動[80]:
![{\displaystyle {\begin{aligned}{\frac {dx}{dt}}&=y+\phi (x)-z+I,\\{\frac {dy}{dt}}&=\psi (x)-y,\\{\frac {dz}{dt}}&=r[s(x-x_{R})-z],\end{aligned}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/d09b646dc48b5bbd1f1f5bed50f9b05985e3d90b)
當中

係指時間, 係指細胞膜電位, 同 係模擬離子通過離子門嘅流動,而 、、、、、、 同 係一啲數值恆定嘅參數,每個呢啲參數都代表咗個系統嘅某一個特性。實驗顯示,呢個模型成功噉以高嘅準確度預測真嘅神經細胞嘅行為[80]。











除咗單一神經細胞模型之外,仲有所謂嘅神經群模型(neural mass model)。顧名思義,神經群模型係指模擬一大柞神經細胞嘅活動嘅模型。呢啲模型會將柞神經細胞當做一個空間上連續嘅神經網絡噉嚟睇,並且喺模擬每一個變數嗰陣,都用嗰個變數橫跨成個網絡嘅平均值嚟計數。呢種做法類似統計力學(statistical mechanics)上嘅研究方法:統計物理學好多時要研究大規模而且有大量組成部件嘅系統(例如係一團普通嘅氣體,入面閒閒地有幾百億粒粒子),所以佢哋用嗰啲數學模型喺研究好多時都有大量神經細胞嘅神經系統嗰陣都有用[81][82]。
藏本模型
[編輯]藏本模型(Kuramoto model;取自日本物理學家藏本由紀個名)係另一個對研究神經振盪同同步化嚟講好有用嘅模型[83][84]。藏本模型會用以角度計嘅相位(phase;指個振盪緊嘅系統處於佢個週期嘅邊一點,例如 0° 代表週期嘅開始點,180° 代表週期嘅一半)嚟代表研究緊嘅神經系統當中某部份嘅活動,忽視嗰個活動嘅波幅[85],並且用(例如)以下呢條式嚟表示啲變數之間嘅關係:
- ;

當中左面嗰嚿表示研究緊嗰橛嘅相位隨時間嘅變化(可以睇導數), 代表個系統嘅自然頻率(natural frequency), 代表嗰橛啲神經細胞嘅相配程度(coupling), 代表嗰橛有幾多個振盪子。如果用日常用語講嘅話,呢條式講嘅嘢如下:研究緊嗰個神經系統部份嘅振盪相位隨時間嘅改變率(條式左面嗰嚿)好大程度上由嗰個部份嘅自然頻率話事,又會受到佢同個系統當中嘅其他部份喺振盪相位上嘅差異()所影響。個研究者知道嗮啲參數嘅數值之後,就可以模擬成個系統嘅行為[84][85]。




藏本模型俾神經科學家廣泛噉攞嚟研究腦入面嗰啲神經振盪活動,而且仲出咗第啲版本嚟更仔細噉提高個模型喺生物學上嘅可信性,例如係有啲模型會考慮埋塊大腦皮層嘅連繫性(即係加多個參數表示「大腦皮層嘅連繫性」)[86]。跟手啲神經科學家仲會攞實際嘅數據嚟驗證呢啲模型[87]。
功能
[編輯]神經振盪對於一隻動物嘅生存嚟講相當重要:神經振盪有份調制多種嘅認知過程,對於神經整合(neural binding;將神經系統嘅唔同資訊結合做一個整體嘅體驗)[88]、神經細胞之間嘅通訊[14]、仲有係運動控制上嘅協調嚟講都好緊要,例如係運動控制(motor control;指一隻動物控制自己身體郁動)上嘅協調噉,一隻動物嘅神經系統一定要能夠無時無刻向身體嘅每嚿肌肉傳適當嘅訊號,先至可以成功噉幫隻動物喺自己嘅生存環境嗰度郁動[17]。
根據廿一世紀初嘅神經科學,腦電波嘅主要功能有以下呢啲:
控制心跳
[編輯]- 控制心跳:神經振盪有幫手調控一個生存必要嘅功能-心跳;要維持一隻動物生存,心跳呢家嘢必需要有節奏,而神經振盪定義上就係神經系統當中一啲有節奏性(有特定頻率同週期)嘅活動,所以理論上經已有可能俾神經系統攞嚟做調節心跳嘅工具(例:想像以頻率恆定嘅腦電波作為射去心臟肌肉嘅訊號);起搏細胞(pacemaker cell)係一種集中喺竇房結(sinoatrial node;右心房裏面一橛)嘅細胞;喺冇乜嘢神經訊號或者荷爾蒙搞佢哋嗰時,起搏細胞會以稍為快過心臟第啲部份嘅細胞嘅速度產生動作電位,會每分鐘自發噉去極化大約 100 次,令到心臟整體喺竇房結嘅引領之下跟住跳動,將血泵去全身嘅各個器官嗰度。除此之外,竇房結亦都同自主神經系統(autonomic nervous system)有好強嘅神經連繫,而自主神經系統會喺有需要嘅時候透過向起搏細胞(例如係以特定頻率)射訊號嚟控制心跳,即係例如係喺要做劇烈運動嗰陣要起搏細胞射動作電位射得快啲,令心跳加速[89]。

感知
[編輯]- 視知覺(visual perception):證據指,腦電波同視覺系統(visual system;指專門由眼嗰度收訊號,處理視覺相關資訊嘅腦區)有相當嘅關係;喺廿世紀尾,有實驗試過搵貓仔嚟研究呢個問題-喺呢個實驗當中,一班貓仔俾研究者開刀,植入咗啲電極落佢哋嘅視覺皮層(visual cortex;大腦皮層當中專門處理視覺資訊嗰一忽)嗰度,用嚟量度佢哋醒緊嗰陣時嘅視覺皮層腦電活動;結果顯示,每當啲貓仔感知到視覺資訊(睇到某啲物件)嗰陣,唔同群嘅神經細胞會進入同步嘅振盪活動;呢啲振盪嘅頻率有成 40 Hz,同一般狀態之下嘅活動好唔一樣,顯示呢啲振盪嘅同步係因為神經細胞之間嘅互動而起嘅[90];第啲研究亦都得到類似嘅結果[91]。
- 第啲感官:除咗視覺之外,亦有神經科學家提議話神經振盪仲喺時間感(chronoception;指人對「過咗幾耐時間」嘅感知)[94]同埋體感(somesthetic sense;指對皮膚表面同體內嘅資訊嘅感知)[95]上擔當咗重要嘅角色。但呢啲假說到咗廿一世紀都仲係一種猜想[96]。
第啲功能
[編輯]- 運動控制(motor control):個腦嘅運動系統入面有好多神經振盪,而呢啲振盪同身體郁動好有關;例如
- 記憶(memory):記憶指一個認知系統儲起資訊嘅能力。神經振盪對於記憶功能嚟講好緊要,尤其係 theta 波:
- 用嚙齒目動物做嘅研究顯示,theta 波喺一隻動物做學習同記憶提取嗰陣會喺海馬體(hippocampus)同內嗅皮質(entorhinal cortex)呢兩個腦區嗰度有零舍大波幅嘅活動,而呢種腦活動俾科學界相信係對長期增強作用(long-term potentiation)嘅過程嚟講不可或缺;而且 theta 波同 gamma 波之間嘅相配似乎係某啲記憶功能-包括情節記憶(episodic memory;指記事嘅記憶,相對於記概念嘅語義記憶)-嘅必要條件[103][104];
- 喺人當中,單一神經細胞射嘅動作電位同區域性嘅 theta 振盪之間嘅協調能夠用嚟預測個人會唔會成功噉記到嘢[105]。
... 等等。
醫療用途
[編輯]喺神經內科(neurology;處理神經系統相關疾病嘅醫學子領域)上,腦電波可以用嚟診斷好多病:
- 顫抖(tremor):指不由自主但有些少節奏性嘅肌肉收縮同放鬆,會令身體某啲部份來回噉郁;顫抖係咁多種不隨意嘅身體郁動當中最常見嘅,可以影響一個人嘅手掌、手臂、眼、面、頭、聲帶、軀幹同埋腳,尤其係以手掌最常見,可以做成生活上極大嘅不便,例如個病人隻手係噉震,搞到佢寫起字上嚟亂嗮大籠。顫抖可以係好多神經系統疾病嘅徵狀,例如柏金遜症(Parkinson's disease)就出咗名會搞到患者多個身體部份出現顫抖,而研究表明咗,柏金遜症患者嘅腦嘅某啲異常腦電波頻率能夠預測個病人顫抖嘅頻率[106]。
-
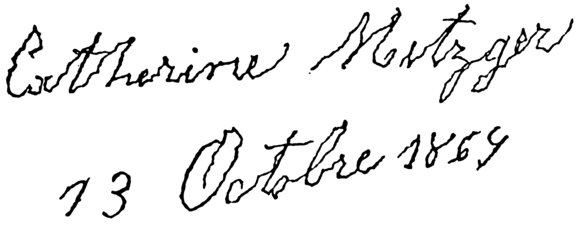 一個柏金遜症患者手寫嘅字;
一個柏金遜症患者手寫嘅字;
由啲筆劃嗰度睇得出佢隻手有節奏性噉震。

- 發羊吊(epilepsy):係一種神經內科病,指病人嘅腦某啲區域有唔正常嘅極高波幅或者極高同步性腦電波,而病人喺病發嗰陣最嚴重會出現成身抽筋嘅情況[107];廿一世紀初有一啲用老鼠同大家鼠做受試者嘅研究發現,好多個體喺受過某啲腦創傷之後會出現發羊吊嘅情況,而呢啲動物嘅腦嘅 theta 波波幅能夠預測好幾種唔同嘅創傷後發羊吊嘅徵狀,所以做呢份研究嗰班科學家提議,話呢個發現可以幫醫護人員控制發羊吊嘅病情-例如係喺個病人嘅腦裝電極,特登壓抑某啲 theta 波活動[108]。

- 過度活躍症(hyperactivity disorder,簡稱「ADHD」):一種搞到病人專注力差嘅病;有研究指,過度活躍症嘅患者喺某啲情況下塊前額葉(prefrontal lobe;大腦皮層近額頭嗰忽)嗰度 theta 波等嘅低頻振盪波幅會大咗;前額葉對於自控等嘅功能嚟講係好緊要嘅,所以「ADHD 病人前額葉有振幅零舍大嘅低頻活動」呢一個觀察一般都俾人認為係表示 ADHD 病人嘅前額葉冇乜好勁嘅活動,反映呢啲患者唔多能夠集中精神控制自己[109][110]。
... 等等。
註釋
[編輯]- ↑ 數值以微伏特(μV)計,同一般家用電器所涉及嘅電壓(100 - 200 V)比起嚟好微弱。
- ↑ 喺物理學上,振盪(oscillation)定義上係指「一個物理量數值喺 、0 同 之間週期性噉上上落落」,當中 係一個大致恆定嘅數值(振幅)。喺腦電波當中,「神經訊號造成嘅微電壓」會有噉嘅上落-所以腦電波可以諗做「神經活動造成嘅振盪」。
- ↑ 入侵性(invasive)腦電量度就係指開刀將電極植入去受試者嘅腦嗰度,用嚟量度個腦深處嘅腦電波。呢種做法會令受試者有相當嘅不適同埋可以造成永久性嘅傷害,喺人身上用會引起道德爭議,所以通常喺人以外嘅動物身上先會用。
- ↑ 即係起碼中層次嘅分析。
- ↑ 但都有啲神經科學家同心理學家唔用呢種分類法。


睇埋
[編輯]文獻
[編輯]- Buzsáki, György (2006). Rhythms of the Brain. Oxford University Press.
- Freeman, Walter (1975). Mass Action in the Nervous System. Academic Press.
- Henry, M. J., & Obleser, J. (2012). Frequency modulation entrains slow neural oscillations and optimizes human listening behavior. Proceedings of the National Academy of Sciences, 109(49), 20095-20100.
- Peelle, J. E., & Davis, M. H. (2012). Neural oscillations carry speech rhythm through to comprehension. Frontiers in psychology, 3, 320.
- Roux, F., & Uhlhaas, P. J. (2014). Working memory and neural oscillations: alpha-gamma versus theta-gamma codes for distinct WM information? (PDF). Trends in cognitive sciences, 18(1), 16-25.
- Ward, L. M. (2003). Synchronous neural oscillations and cognitive processes (PDF). Trends in cognitive sciences, 7(12), 553-559.
攷
[編輯]- ↑ Neural Oscillation. Medical dictionary.
- ↑ Klimesch, W. (1999). EEG alpha and theta oscillations reflect cognitive and memory performance: a review and analysis. Brain Res Brain Res Rev, 29:169–95.
- ↑ Rangaswamy M., Porjesz B., Chorlian D.B., Wang K., Jones K.A., Bauer L.O., Rohrbaugh J., O'Connor S.J., Kuperman S., Reich T., Begleiter (2002). "Beta power in the EEG of alcoholics". Biological Psychology. 52 (8): 831–842.
- ↑ Coenen, Anton; Edward Fine; Oksana Zayachkivska (2014). "Adolf Beck: A Forgotten Pioneer In Electroencephalography". Journal of the History of the Neurosciences. 23 (3): 276–286.
- ↑ Pravdich-Neminsky, VV. (1913). "Ein Versuch der Registrierung der elektrischen Gehirnerscheinungen". Zentralblatt für Physiologie. 27: 951–60.
- ↑ Llinas, R. R. (2014). "Intrinsic electrical properties of mammalian neurons and CNS function: a historical perspective". Front Cell Neurosci. 8: 320.
- ↑ Vansteensel, M. J., Pels, E. G., Bleichner, M. G., Branco, M. P., Denison, T., Freudenburg, Z. V., ... & Van Rijen, P. C. (2016). Fully implanted brain–computer interface in a locked-in patient with ALS. New England Journal of Medicine, 375(21), 2060-2066.
- ↑ Birbaumer, Neils (2006). "Breaking the silence: Brain-computer interfaces (BCI) for communication and motor control". Psychophysiology. 43 (6): 517–32.
- ↑ 9.0 9.1 9.2 Hodgkin, A. L., & Huxley, A. F. (1952). "A quantitative description of membrane current and its application to conduction and excitation in nerve". The Journal of Physiology. 117 (4): 500–544.
- ↑ Fraser, A.; Frey, A. H. (1968). "Electromagnetic emission at micron wavelengths from active nerves". Biophysical Journal. 8 (6): 731–734.
- ↑ Adrian, E. D. (1964). The Basis of Sensation. Hafner, New York.
- ↑ Llinas R., Yarom Y. (1986). "Oscillatory properties of guinea-pig inferior olivary neurones and their pharmacological modulation: an in vitro study". J Physiol. 376: 163&ndash, 182.
- ↑ Niedermeyer E.; da Silva F.L. (2004). Electroencephalography: Basic Principles, Clinical Applications, and Related Fields. Lippincott Williams & Wilkins.
- ↑ 14.0 14.1 Fries P. (2005). "A mechanism for cognitive dynamics: neuronal communication through neuronal coherence". TICS. 9 (10): 474–480.
- ↑ Fell J., Axmacher N. (2011). "The role of phase synchronization in memory processes". Nature Reviews Neuroscience. 12 (2): 105–118.
- ↑ 16.0 16.1 16.2 Haken H. (1996). Principles of brain functioning. Springer.
- ↑ 17.0 17.1 Schnitzler A., Gross J. (2005). "Normal and pathological oscillatory communication in the brain". Nature Reviews Neuroscience. 6 (4): 285, 296.
- ↑ 18.0 18.1 18.2 Izhikevich E.M. (2007). Dynamical systems in neuroscience. Cambridge, Massachusetts: The MIT Press.
- ↑ Burrow T. (1943). "The neurodynamics of behavior. A phylobiological foreword". Philosophy of Science. 10 (4): 271–288.
- ↑ Aidley D.J., Stanfield P.R. (1996). Ion Channels: Molecules in Action. Cambridge: Cambridge University Press.
- ↑ 21.0 21.1 Wang X.J. (2010). "Neurophysiological and computational principles of cortical rhythms in cognition". Physiol Rev. 90 (3): 1195–1268.
- ↑ Neuron action potentials: The creation of a brain signal. Khan Academy.
- ↑ Lagercrantz, Hugo; Hanson, M. A.; Ment, Laura R.; Peebles, Donald M., eds. (7 January 2010). The Newborn Brain: Neuroscience and Clinical Applications. Cambridge University Press. p. 3.
- ↑ Nunez P.L., Srinivasan R. (1981). Electric fields of the brain: The neurophysics of EEG. Oxford University Press.
- ↑ Zeitler M., Daffertshofer A., Gielen C.C. (2009). "Asymmetry in pulse-coupled oscillators with delay". Phys. Rev. E. 79 (6): 065203(R).
- ↑ Cardin J.A., Carlen M., Meletis K., Knoblich, U., Zhang F., Deisseroth K., Tsai L.H., Moore C.I. (2009). "Driving fast-spiking cells induces gamma rhythm and controls sensory responses". Nature. 459 (7247): 663–U63.
- ↑ Llinás R., Ribary U., Contreras D., Pedroarena C. (November 1998). "The neuronal basis for consciousness". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 353 (1377): 1841–9.
- ↑ Bollimunta A., Mo J., Schroeder C.E., Ding M. (March 2011). "Neuronal mechanisms and attentional modulation of corticothalamic α oscillations". The Journal of Neuroscience. 31 (13): 4935–43.
- ↑ Suffczynski P., Kalitzin S., Pfurtscheller G., Lopes da Silva F.H. (December 2001). "Computational model of thalamo-cortical networks: dynamical control of alpha rhythms in relation to focal attention". International Journal of Psychophysiology. 43 (1): 25–40.
- ↑ Albert, B., Zhang, J., Noyvirt, A., Setchi, R., Sjaaheim, H., Velikova, S., & Strisland, F. (2016, July). Automatic EEG processing for the early diagnosis of traumatic brain injury. In 2016 World Automation Congress (WAC) (pp. 1-6). IEEE.
- ↑ Makeig, S. (1993). Auditory event-related dynamics of the EEG spectrum and effects of exposure to tones. Electroencephalogr Clin Neurophysiol, 86:283–93.
- ↑ 32.0 32.1 Ward, L. M. (2003). Synchronous neural oscillations and cognitive processes. Trends in cognitive sciences, 7(12), 553-559.
- ↑ Nunez, P. L. (1974). The brain wave equation: a model for the EEG. Mathematical Biosciences, 21(3-4), 279-297.
- ↑ Bracewell R., Kahn P.B.. The Fourier transform and its applications. Am J Phys. 1966;34:712.
- ↑ Gerrard P., Malcolm R. (June 2007). "Mechanisms of modafinil: A review of current research". Neuropsychiatr Dis Treat. 3: 349–64.
- ↑ Berger H., Gray C.M. (1929). "Uber das Elektroenkephalogramm des Menschen". Arch Psychiat Nervenkr. 87: 527–570.
- ↑ Dement W., Kleitman N. (1957). "Cyclic variations in EEG during sleep and their relation to eye movements, body motility and dreaming". Electroencephalogr Clin Neurophysiol. 9 (4): 673–90.
- ↑ Engel A.K., Singer W. (2001). "Temporal binding and the neural correlates of sensory awareness". Trends in Cognitive Sciences. 5 (1): 16–25.
- ↑ 39.0 39.1 Varela F., Lachaux J.P., Rodriguez E., Martinerie J. (2001). "The brainweb: phase synchronization and large-scale integration". Nature Reviews Neuroscience. 2 (4): 229–239.
- ↑ WHAT ARE BRAINWAVES?. brainworks.
- ↑ Dang-Vu, T. T., Desseilles, M., Laureys, S., Degueldre, C., Perrin, F., Phillips, C., ... & Peigneux, P. (2005). Cerebral correlates of delta waves during non-REM sleep revisited. Neuroimage, 28(1), 14-21.
- ↑ 42.0 42.1 Understanding the brainwaves of your children 互聯網檔案館嘅歸檔,歸檔日期2020年3月28號,..
- ↑ 43.0 43.1 Kirmizi-Alsan, Elif; Bayraktaroglu, Zubeyir; Gurvit, Hakan; Keskin, Yasemin H.; Emre, Murat; Demiralp, Tamer (2006). "Comparative analysis of event-related potentials during Go/NoGo and CPT: Decomposition of electrophysiological markers of response inhibition and sustained attention". Brain Research. 1104 (1): 114–28.
- ↑ Gloor, P., Ball, G., & Schaul, N. (1977). Brain lesions that produce delta waves in the EEG. Neurology, 27(4), 326-326.
- ↑ Hirose, G., Saeki, M., Kosoegawa, H., Takado, M., Yamamoto, T., & Tada, A. (1981). Delta waves in the EEGs of patients with intracerebral hemorrhage. Archives of neurology, 38(3), 170-175.
- ↑ Clarke, A. R., Barry, R. J., McCarthy, R., & Selikowitz, M. (1998). EEG analysis in attention-deficit/hyperactivity disorder: a comparative study of two subtypes. Psychiatry research, 81(1), 19-29.
- ↑ Park, J., Xu, L., Sridhar, V., Chi, M., & Cauwenberghs, G. (2011, August). Wireless dry EEG for drowsiness detection. In 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society (pp. 3298-3301). IEEE.
- ↑ Kurt, M. B., Sezgin, N., Akin, M., Kirbas, G., & Bayram, M. (2009). The ANN-based computing of drowsy level. Expert Systems with Applications, 36(2), 2534-2542.
- ↑ Cahn, B. Rael; Polich, John (2006). Meditation states and traits: EEG, ERP, and neuroimaging studies. Psychological Bulletin. 132 (2): 180–211.
- ↑ Kolev V., Başar-Eroglu C., Aksu F., Başar E. (April 1994). "EEG rhythmicities evoked by visual stimuli in three-year-old children". Int. J. Neurosci. 75: 257–70.
- ↑ Gargiulo, G., Bifulco, P., Calvo, R. A., Cesarelli, M., Jin, C., & van Schaik, A. (2008, November). A mobile EEG system with dry electrodes. In Biomedical Circuits and Systems Conference, 2008. BioCAS 2008. IEEE (pp. 273-276). IEEE.
- ↑ Chokroverty, S. (1975). “Alpha‐like” rhythms in electroencephalograms in coma after cardiac arrest. Neurology, 25(7), 655-655.
- ↑ Baumeister J., Barthel T., Geiss K.R., Weiss M. (2008). "Influence of phosphatidylserine on cognitive performance and cortical activity after induced stress". NUTRITIONAL NEUROSCIENCE. 11 (3): 103–110.
- ↑ Frohlich, Joel; Senturk, Damla; Saravanapandian, Vidya; Golshani, Peyman; Reiter, Lawrence; Sankar, Raman; Thibert, Ronald; DiStefano, Charlotte; Cook, Edwin; Jeste, Shafali (December 2016). "A Quantitative Electrophysiological Biomarker of Duplication 15q11.2-q13.1 Syndrome". PLOS One. 11: e0167179.
- ↑ Kisley, Michael A.; Cornwell, Zoe M. (2006). "Gamma and beta neural activity evoked during a sensory gating paradigm: Effects of auditory, somatosensory and cross-modal stimulation". Clinical Neurophysiology. 117 (11): 2549–63.
- ↑ Kanayama, Noriaki; Sato, Atsushi; Ohira, Hideki (2007). "Crossmodal effect with rubber hand illusion and gamma-band activity". Psychophysiology. 44 (3): 392–402.
- ↑ Jensen, O., Kaiser, J., & Lachaux, J. P. (2007). Human gamma-frequency oscillations associated with attention and memory. Trends in neurosciences, 30(7), 317-324.
- ↑ Başar, E. (2013). A review of gamma oscillations in healthy subjects and in cognitive impairment. International Journal of Psychophysiology, 90(2), 99-117.
- ↑ Gastaut, H. (1952). "Electrocorticographic study of the reactivity of rolandic rhythm". Revue neurologique. 87 (2): 176–82.
- ↑ Oberman, Lindsay M.; Hubbard, Edward M.; McCleery, Joseph P.; Altschuler, Eric L.; Ramachandran, Vilayanur S.; Pineda, Jaime A. (2005). "EEG evidence for mirror neuron dysfunction in autism spectrum disorders". Cognitive Brain Research. 24 (2): 190–8.
- ↑ Ibáñez, Vanessa; Silva, Josep; Cauli, Omar (2018-05-25). "A survey on sleep assessment methods". PeerJ. 6: e4849.
- ↑ Jäntti, V., & Sloan, T. B. (2008). EEG and anesthetic effects. Handbook of Clinical Neurophysiology, 8, 77-93.
- ↑ Recommendations for the Practice of Clinical Neurophysiology: Guidelines of the International Federation of Clinical Physiology (EEG Suppl. 52) Editors: G. Deuschl and A. Eisen q 1999 International Federation of Clinical Neurophysiology. All rights reserved. Published by Elsevier Science B.V.
- ↑ Pfurtscheller G.; da Silva F.H.L. (1999). "Event-related EEG/MEG synchronization and desynchronization: basic principles". Clin Neurophysiol. 110 (11): 1842&ndash, 1857.
- ↑ Freyer F., Aquino K., Robinson P.A., Ritter P., Breakspear M. (2009). "Bistability and non-Gaussian fluctuations in spontaneous cortical activity". J Neurosci. 29 (26): 8512–8524.
- ↑ Fox M.D., Raichle M.E. (2007). "Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging". Nat Rev Neurosci. 8 (9): 700&ndash, 711.
- ↑ Laufs H., Kraków K., Sterzer P., Eger E., Beyerle A., Salek-Haddadi A., Kleinschmidt A. (2003). "Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging". PNAS. 100 (19): 11053–11058.
- ↑ Mathewson K.E., Gratton G., Fabiani M., Beck D.M., Ro T. (2009). "To see or not to see: Prestimulus α phase predicts visual awareness". J Neurosci. 29 (9): 2725–32.
- ↑ Busch N.A., Dubois J., VanRullen R. (2009). "The phase of ongoing EEG oscillations predicts visual perception". J Neurosci. 29 (24): 7869–76.
- ↑ van Dijk H., Schoffelen J.M., Oostenveld R., Jensen O. (2008). "Prestimulus oscillatory activity in the alpha band predicts visual discrimination ability". J Neurosci. 28 (8): 1816–1823.
- ↑ Kandel, E.; Schwartz, J.; Jessel, T.M. (1991). Principles of Neural Science (3rd ed.). Elsevier.
- ↑ Diedrich, F. J., & Warren Jr, W. H. (1995). Why change gaits? Dynamics of the walk-run transition. Journal of Experimental Psychology: Human Perception and Performance, 21(1), 183.
- ↑ Tallon-Baudry C., Bertrand O. (1999). "Oscillatory gamma activity in humans and its role in object representation". Trends Cogn Sci. 3 (4): 151&ndash, 162.
- ↑ Tass P.A. (2007). Phase resetting in medicine and biology: stochastic modelling and data analysis. Berlin Heidelberg: Springer-Verlag.
- ↑ Pikovsky A., Rosenblum M., Kurths J. (2001). Synchronization: a universal concept in nonlinear sciences. Cambridge University Press.
- ↑ Barry, R. J. (2009). Evoked activity and EEG phase resetting in the genesis of auditory Go/NoGo ERPs. Biological psychology, 80(3), 292-299.
- ↑ Makeig S., Westerfield M., Jung T.P., Enghoff S., Townsend J., Courchesne E., Sejnowski T.J. (2002). "Dynamic brain sources of visual evoked responses". Science. 295 (5555): 690&ndash, 694.
- ↑ Buzsaki G., Draguhn A. (2004). "Neuronal oscillations in cortical networks". Science. 304 (5679): 1926–1929.
- ↑ Whittington M.A., Traub R.D., Kopell N., Ermentrout B., Buhl E.H. (2000). "Inhibition-based rhythms: experimental and mathematical observations on network dynamics". Int J Psychophysiol. 38 (3): 315–336.
- ↑ 80.0 80.1 Hindmarsh J. L.; Rose R. M. (1984). "A model of neuronal bursting using three coupled first order differential equations". Proc. R. Soc. London, Ser. B. 221: 87–102.
- ↑ Wendling F., Bellanger J.J., Bartolomei F., Chauvel P. (2000). "Relevance of nonlinear lumped-parameter models in the analysis of depth-EEG epileptic signals". Biol Cybern. 83 (4): 367–378.
- ↑ Bressloff P.C., Cowan J.D. (2003) Spontaneous pattern formation in primary visual cortex. In: J. Hogan, A.R. Krauskopf, M. di Bernado, R.E. Wilson (Eds.), Nonlinear dynamics and chaos: where do we go from here?.
- ↑ Acebrón, J. A., Bonilla, L. L., Vicente, C. J. P., Ritort, F., & Spigler, R. (2005). The Kuramoto model: A simple paradigm for synchronization phenomena. Reviews of modern physics, 77(1), 137.
- ↑ 84.0 84.1 Kuramoto Y. (1984). Chemical Oscillations, Waves, and Turbulence. Dover Publications.
- ↑ 85.0 85.1 Ermentrout B. (1994). "An introduction to neural oscillators". In F Ventriglia. Neural Modeling and Neural Networks. pp. 79–110.
- ↑ Breakspear M., Heitmann S., Daffertshofer A. (2010). "Generative models of cortical oscillations: Neurobiological implications of the Kuramoto model". Front Hum Neurosci. 4: 190.
- ↑ Cabral J., Hugues E., Sporns O., Deco G. (2011). "Role of local network oscillations in resting-state functional connectivity". NeuroImage. 57 (1): 130–9.
- ↑ 88.0 88.1 Singer W. (1993). "Synchronization of cortical activity and its putative role in information processing and learning". Annu Rev Physiol. 55: 349–374.
- ↑ Neil A. Campbell ... [et al.] (2006). Biology : concepts & connections (5th ed.). San Francisco: Pearson/Benjamin Cummings. p. 473.
- ↑ 90.0 90.1 Gray C.M., König P., Engel A.K., Singer W. (1989). "Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties". Nature. 338 (6213): 334–337.
- ↑ Eckhorn R., Bauer R., Jordan W., Brosch M., Kruse W., Munk M., Reitboeck H.J. (1988). "Coherent oscillations: A mechanism of feature linking in the visual cortex? Multiple electrode and correlation analyses in the cat". Biol Cybern. 60 (2): 121–130.
- ↑ Milner P.M. (1974). "A model for visual shape recognition". Psychol. Rev. 81 (6): 521–535.
- ↑ Singer W., Gray C.M. (1995). "Visual feature integration and the temporal correlation hypothesis". Annu Rev Neurosci. 18: 555–586.
- ↑ Buhusi C.V., Meck W.H. (2005). "What makes us tick? Functional and neural mechanisms of interval timing". Nature Reviews Neuroscience. 6 (10): 755–65.
- ↑ Ahissar E., Zacksenhouse M. (2001). "Temporal and spatial coding in the rat vibrissal system". Prog Brain Res. Progress in Brain Research. 130: 75–87.
- ↑ Burns S.P., Xing D., Shapley R.M. (2011). "Is gamma-band activity in the local field potential of V1 cortex a "clock" or filtered noise?". J Neurosci. 31 (26): 9658–9664.
- ↑ Pfurtscheller G., Aranibar A. (1977). "Event-related cortical desynchronization detected by power measurements of scalp EEG". Electroencephalogr Clin Neurophysiol. 42 (6): 817–826.
- ↑ Conway, B.A.; Halliday, D.M.; Farmer, S.F. (1995). "Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man". J Physiol. 489 (3): 917–924.
- ↑ Murthy V.N., Fetz E.E. (1996). "Oscillatory activity in sensorimotor cortex of awake monkeys: Synchronization of local field potentials and relation to behavior". J Neurophysiol. 76 (6): 3949–3967.
- ↑ Sanes J.N., Donoghue J.P. (1993). "Oscillations in local-field potentials of the primate motor cortex during voluntary movement". PNAS. 90 (10): 4470–4474.
- ↑ Salenius S., Portin K., Kajola M., et al. (1997). "Cortical control of human motoneuron firing during isometric contraction". J Neurophysiol. 77 (6): 3401–3405.
- ↑ Baker S.N., Olivier E., Lemon R.N. (1997). "Coherent oscillations in monkey motor cortex and hand muscle EMG show task-dependent modulation". J Physiol. 501 (1): 225–241.
- ↑ Buszaki G. (2006). Rhythms of the brain. Oxford University Press.
- ↑ Nyhus E., Curran T. (June 2010). "Functional role of gamma and theta oscillations in episodic memory". Neuroscience and Biobehavioral Reviews. 34 (7): 1023–35.
- ↑ Rutishauser U., Ross I.B., Mamelak A.N., Schuman E.M. (April 2010). "Human memory strength is predicted by theta-frequency phase-locking of single neurons". Nature. 464 (7290): 903–7.
- ↑ McAuley J.H., Marsden C.D. (2000). "Physiological and pathological tremors and rhythmic central motor control". Brain. 123 (8): 1545–1567.
- ↑ V. Shusterman and W. C. Troy. From baseline to epileptiform activity: A path to synchronized rhythmicity in large-scale neural networks. Phys Rev E Stat Nonlin Soft Matter Phys. 2008;77(6 Pt 1):061911.
- ↑ Milikovsky, D. Z., Weissberg, I., Solomon-Kamintsky, L., Lippmann, K., Schefenbauer, O., Frigerio, F., ... & Vezzani, A. (2017). Electrocorticographic dynamics as a novel biomarker in five models of epileptogenesis. Journal of Neuroscience, 2446-16.
- ↑ Ogrim, G., Kropotov, J., & Hestad, K. (2012). The quantitative EEG theta/beta ratio in attention deficit/hyperactivity disorder and normal controls: sensitivity, specificity, and behavioral correlates. Psychiatry research, 198(3), 482-488.
- ↑ Putman, P., van Peer, J., Maimari, I., & van der Werff, S. (2010). EEG theta/beta ratio in relation to fear-modulated response-inhibition, attentional control, and affective traits. Biological psychology, 83(2), 73-78.
